
Les articles Ouest-Paléo

Avant la médiatisation des découvertes fossiles du gisement d'Angeac-Charente un autre gisement charentais a attiré les chercheurs du monde entier, qu'ils soient professionnels ou amateurs, le gisement de Cherves de Cognac.
Le gisement de Cherves de Cognac est daté d'une époque où les archives fossiles sont rares, le Berriasien. La datation de ce gisement le rendait déjà très intéressant, mais la mise à jour d'une faune exceptionnelle rendit encore plus important ce gisement.
En effet Cherves de Cognac a livré une faune remarquable par sa diversité et sa richesse : reptiles, poissons, mammifères, amphibiens, gastéropodes, bivalves…
 La carrière de Chamblanc à différentes époques.
La carrière de Chamblanc à différentes époques.
Contexte géographique et stratigraphique.
Le gisement de Cherves de Cognac est situé sur la commune de Cherves-Richemont, à une dizaine de kilomètres au Nord-est de Cognac (Charente, France). Cherves-Richemont est née de la fusion de Cherves de Cognac et de Richemont, le gisement est situé sur l’ancienne commune de Cherves de Cognac et il a été décidé d’utiliser ce nom pour toute mention concernant le gisement, compte tenu de la notoriété du Cognac sur le plan international.
Il se trouve dans une carrière de gypse de l’entreprise Garandeau, carrière en activité et interdite au public. Le gypse, matière première pour la fabrication du plâtre, est exploité dans la région depuis l’Antiquité et représentait en 1980 5% de la production nationale. Cette carrière produit en moyenne 420 000 tonnes de gypse par an.
Cette carrière est située dans le Bassin aquitain, à la limite des terrains du Jurassique supérieur (Tithonien, -150 à -145 millions d’années) et du Crétacé inférieur (Berriasien, -145 à -139 millions d’années).
Il s’agit d’une carrière où est exploité le gypse qui, après cuisson et broyage, est transformé en plâtre.
Outre le gypse on trouve dans cette carrière des marnes, du calcaire et des argiles noires. Ces couches sont peu fossilifères, sauf quelques unes qui ont livrée une faune exceptionnelle et qui forment un bone-bed.
A l’origine les sédiments ont été datés du Purbeckien, mais de nouvelles études leur ont attribués un âge légèrement plus récent, Berriasien inférieur à moyen (-141 à -135 millions d’années), études basées sur l’analyse de certaines caractéristiques : ostracodes, charophytes et dinoflagellés.
 Les niveaux à tétrapodes de la coupe de Cherves de Cognac.
Les niveaux à tétrapodes de la coupe de Cherves de Cognac.
(A) Coupe globale, avec ses trois parties majeures. (B) agrandissement sur les niveaux à macro-restes de tétrapodes. (C) Photographie des niveaux à macro-restes de tétrapodes, les limites des niveaux sont indiqués en trait fins (les tirets expriment une incertitude sur la validité du niveau C34), le trait épais matérialise le niveau C36, présent occasionnellement, qui recoupe les niveaux sous-jacents. A et B, modifié d’après El Albani et al.(2003). Echelles (A) et (B), 1 m ; (C), 50cm.
In BILLON-BRUYAT Jean-Paul, 2003: Les écosystèmes margino-littoraux du Jurassique terminal et du Crétacé basal d'Europe occidentale: biodiversité, biogéochimie et l'événement biotique de la limite Jurassique-Crétacé
Historique des fouilles
Si des fouilles ont été entreprises au début des années 2000 par JM Mazin, les fossiles de Cherves sont connus depuis bien plus longtemps.
Henri Coquand (1813-1881), géologue et paléontologue charentais, en fait mention en 1862 (Description physique, géologique, paléontologique et minéralogique du département de la Charente. Tome deuxième), il dresse une liste de différents fossiles rencontrés dans les argiles gypsifères de la région de Cognac : dents et écailles de Lepidotus, 5 espèces de gastéropodes et 7 espèces de bivalves. Les couches sont datées à l’époque du Purbeckien, mais Henri Coquand ne précise pas le niveau de découverte de ces fossiles.
Quelques années plus tard (1875), alors qu’il est professeur de géologie à Marseille, Henri Coquand décrira l’étage du Berriasien (à Berrias, en Ardèche). De nos jours les marnes de Cherves-de-Cognac sont datées du Berriasien.
Il faut attendre la fin du XXème siècle pour que le gisement refasse surface : Guides géologiques régionaux : Poitou, Vendée, Charentes, Gabilly & al, 1978 ; notice carte géologique, Bourgueil & al, 1986 ; Buffetaut et al, Poissons et reptiles continentaux du Purbeckien de la région de Cognac (Charente), 1989.
Malgré cela le gisement de Cherves reste encore peu connu et il faudra attendre les années 90 pour qu’il se fasse enfin connaître du grand public grâce à la persévérance de Thierry Lenglet, amateur charentais, qui finit par convaincre les autorités scientifiques de l’intérêt du site, site qui va devenir l’un des plus importants pour le Berriasien.
Et c’est ainsi qu’en 2001 Jean-François Tournepiche, conservateur du Musée d’Angoulême, et Jean-Michel Mazin, professeur à l’université de Poitiers, dirigent une première fouille test pour évaluer l’intérêt du gisement.
En 2 semaines ces fouilles dépasseront toutes leurs attentes, que soit au niveau de la richesse ou de la diversité des restes fossiles : crocodiles, tortues, requins, poissons, dinosaures, ptérosaures… Ainsi près de 150 ossements seront découverts, dont de nombreux restes de crocodylomorphes, notamment plusieurs crânes complets de Goniopholis, et le tamisage révélera un potentiel très élevé.
S’ensuivront 6 années de fouilles où de nombreuses découvertes seront effectuées.
La 1ère campagne de fouille, qui réunira outre les nombreux fouilleurs, une vingtaine de spécialistes internationaux, permettra la découverte d’environ 400 spécimens, représentant une cinquantaine d’espèces, dont des mollusques, crustacés, poissons, tortues, requins, dinosaures, ptérosaures, mammifères… La plupart des fossiles découverts ne sont que des ossements isolés.
Les sédiments ont été lavés et tamisés, révélant des milliers d’autres fossiles, notamment des dents infra-millimétriques, des coquilles d’œufs de reptiles…
Les fouilles suivantes ont permis d’enrichir ces découvertes, notamment grâce à la mise en place d’une station de tamisage permettant de traiter plus de 30 tonnes de sédiments. Elles ont aussi permis de suivre l’évolution du site au Berriasien, en fouillant couche par couche les chercheurs ont ainsi pu avoir une vision plus globale de la faune et de l’environnement.
Au 27 juin 2008, 1792 macro-restes et 42763 micro-restes avaient été mis à jour.
Parmi les découvertes les plus importantes l’on peut citer le Camarasaurus, connu par des os et des dents, des mammifères, connus par de minuscules dents, et un oiseau, connu aussi par une dent.
Ces fouilles ont été possibles grâce au financement des collectivités publiques et du mécénat d’entreprises : groupe Garandeau, Musée d’Angoulême, CNRS, universités de Poitiers et de Lyon, Conseil Général de Charente, communauté de communes de Cognac, commune de Cherves-Richemont, Groupe de Recherche du Musée d’Angoulême (GERMA), association PaleoAquitania…
Outre le travail des équipes du CNRS et des universités de Lyon et Poitiers, ce gisement a vu de nombreuses collaborations scientifiques nationales et internationales : Museum National d’Histoire Naturelle de Paris, Geologisk Museum de Copenhague (Danemark), Natural History Museum de Londres (Grande-Bretagne), University College de Londres (Grande-Bretagne), University de Wolverhampton (Grande-Bretagne), Université de Würzburg (Allemagne), Ecole Polytechnique de Zürich (Suisse), Université de Stockholm (Suède), Université de Barcelone (Espagne)…
 Pholidosaures et stégosaure Cherves by Mazan
Pholidosaures et stégosaure Cherves by Mazan
Contenu fossilifère.
La collecte des micro-restes par tamisage a permis de faire de grandes découvertes, notamment au niveau des dents.
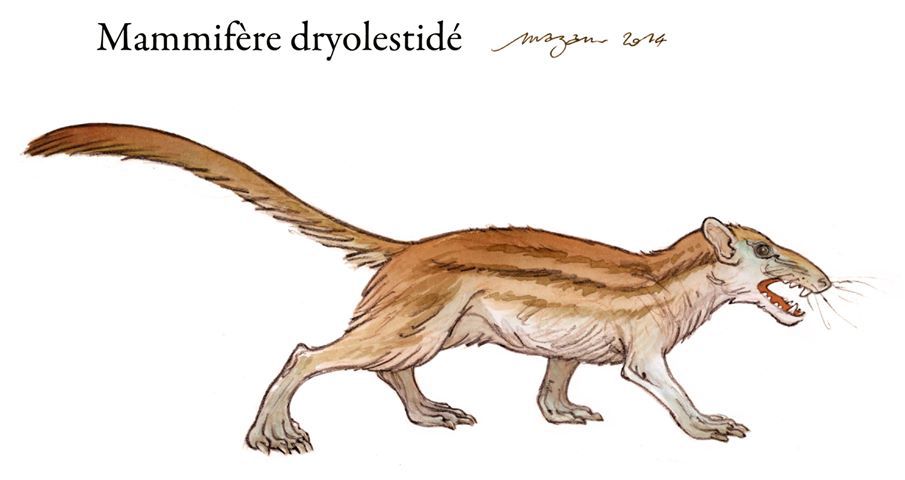
Plusieurs dizaines de milliers de minuscules dents ont ainsi été découvertes, l’on y retrouve du poisson, du requin, du crocodile... Mais certaines sortent du lot, ainsi une cinquantaine appartiennent à de petits mammifères et une à un oiseau.
Ces mammifères, dont les restes sont peu connus, étaient de plusieurs espèces :
-dryolestidae
-Spalacotherium evansae
-spalacotheriidae cf. Spalacotherium
-spalacotheriidae indet.
-triconodontinae indet.
-dryolestidae cf. Laolestes
-dryolestidae indet.
-peramuridae cf Peramus
-Pinheirodon pygmaeus
-Thereuodon sp.
Ces découvertes en font l’un des sites les plus importants pour les mammifères crétacés européens.
Quand à l’oiseau il semblerait qu’il s’agisse d’un archaeopterygidae, la dent étant très proche morphologiquement des dents d’Archaeopteryx du Tithonien allemand et du Kimméridgien portugais.
En plus de ces restes a été retrouvée une faune riche et variée.
Dans les airs on retrouve des pterodactyloidea, ptérosaures à queue courte, et des azhdarchidae (obs. pers.), ptérosaures à grande tête.
Ces reptiles sont connus par des os : fémur, métacarpe, phalange…
 from the Lower Cretaceous of France extends the spatial and temporal occurrence of the earliest birds. Louchart and Pouech 2017")


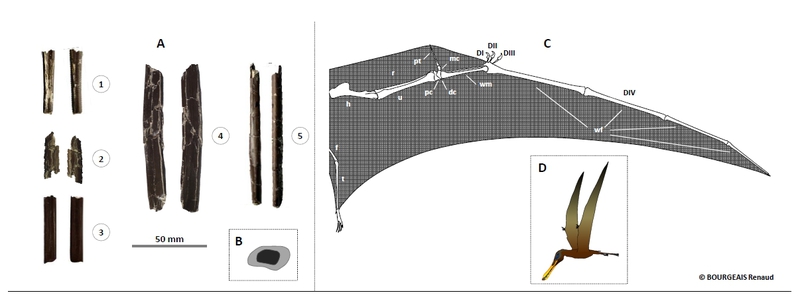



Dans les eaux, les requins et poissons osseux étaient abondants, l’une des espèces les plus retrouvées est Lepidotes (depuis devenu Scheenstia), poisson osseux dont on a découverts écailles, dents, fragments de corps.
Il y avait aussi, pour les chondrichtyens, Parvodus celsucuspus (dents, épines céphaliques et dorsales), Polyacrodus (dents, épines céphaliques et dorsales), Lissodus (dents, épines céphaliques et dorsales), Hybodus (dents, épines céphaliques et dorsales), Belemnobatis variabilis (dents), et pour les ostéichtyens, Belonostomus (dents), Caturus (dents), coelacanthidae, Thrissops (dents).













Les crocodiles et les tortues étaient aussi nombreux et variés.
Pour les crocodiles, pas moins de 7 espèces sont connues :
-Goniopholis simus
-Goniopholis crassidens
-Pholidosaurus purbeckensis
-Bernissartia fagesii
-bernissartidae ind.
-Theriosuchus pusillus
-Theriosuchus nov. sp.
Ils sont connus par de nombreux fossiles, dents, ostéodermes, vertèbres, crânes, coquilles d’œufs…
Pour Goniopholis plus d’une dizaine de crâne ont été découverts, ainsi qu’un squelette complet.
Pour Theriosuchus plusieurs squelettes quasi-complets ont été mis à jour, dont 2 en partie articulés.
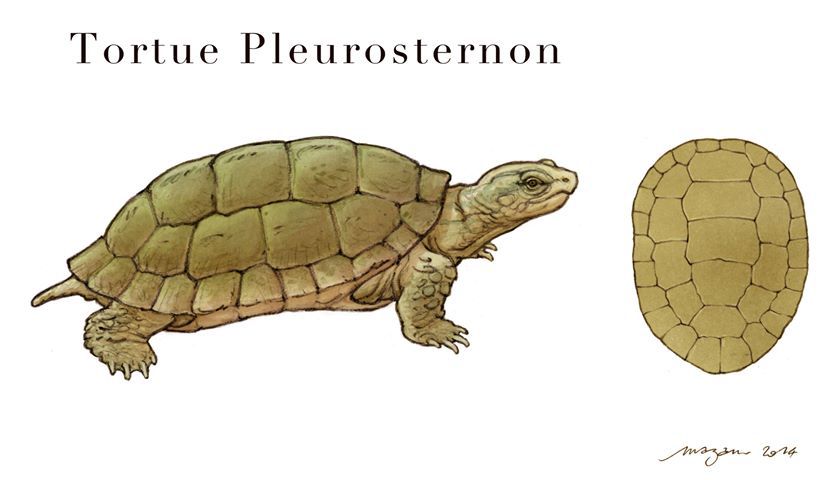
Les tortues, quand à elles, sont connues par 2 espèces :
-Tretosternon
-Pleurosternon
Les restes découverts consistent en de nombreux ostéodermes, des coquilles d’œufs, des os et de rares carapaces plus ou moins complètes.















On a aussi retrouvé de nombreux restes d’amphibiens, fragments de mandibules et humérus d’albanerpetonidae notamment.
Quelques restes de lépidosauriens furent aussi découverts, dont un squelette d’Homeosaurus maximiliani, un sphénodonte, et des fragments de mâchoires de squamates.
 Homoeosaurus cf. maximiliani, Berrisien, Charente
Homoeosaurus cf. maximiliani, Berrisien, Charente
A cela s’ajoute des animaux impressionnants: les dinosaures.
De nombreux restes ont été découverts, allant d’os de monstres de plusieurs dizaines de mètres de longs, à de petites créatures de quelques dizaines de cm.
Le plus impressionnant est sans conteste le sauropode, il s’agit d’une nouvelle espèce de Camarasaurus, non encore décrite à ce jour. Les Camarasaurus, sont des sauropodes de taille moyenne, surtout connu en Amérique du Nord (Dinosaur National Park).
A Cherves pas de squelette complet déterré, mais de beaux restes : dents, vertèbres, os…
A noter qu’un squelette partiel a été découvert par T. Lenglet et confié au CNRS de Lyon pour étude. Squelette comprenant « des éléments de du crâne à la queue, indiquant que le cadavre complet s’est déposé dans le milieu margino-littoral», Billon-Bruyat, 2003.
A ce géant charentais (dépassé depuis par ceux d’Angeac), s’ajoute de plus petites créatures :
-théropodes ind., de taille variées.
-dromaeosauridae.
-titanosauriformes
-Nuthetes destructor.
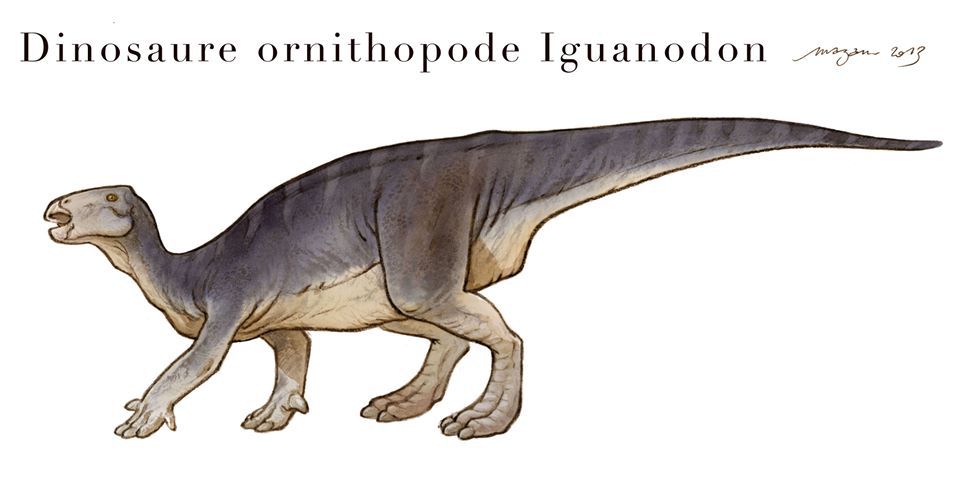
-iguanodontidae.
-ornithopodes indet.
-stegosauridae (Dacentrurus ?), ces restes sont la 2ème occurrence européenne crétacée de stégosaures.
Ces espèces sont surtout connues par des dents isolées et quelques griffes.











On a donc une faune de tétrapodes à composante continentale, avec certains éléments occupant potentiellement le milieu margino-littoral, ce qui confirme l’hypothèse d’un milieu lagunaire, sous influence d’eau douce et marine.
A tous ces vertébrés s’ajoute aussi une riche faune d’invertébrés :
- des ostracodes, minuscules crustacés vivant dans une carapace formée de 2 valves, Cypridea cf. sagena, Macrodentina sp., Cetacella armata, Fabanella boloniensis, Asciocythere sp., Rhinocypris jurassica…
- des charophytes, algues vertes, Porochara kimmeridgiensis, Clavator grovesii…
-des dinoflagellés, protistes, Systematophora palmula, Geiselodinium sp. …
-des gastéropodes, tels que Viviparus ou des hydrobiidae.
-des bivalves, tels que Protocardia intexta ou Corbula inflexa.

 : nouvelles données biostratigraphiques. 2004)")

A tout cela s’ajoute aussi des restes de végétaux, avec notamment un tronc d’Agathoxylon et des pollens (Deltoidospora, Cyathidites, Alisporites, Podocarpites, Eucommidites troedssonii, Vitreisporites pallidus…)
 Agathoxylon, Berriasien. Musée d'Angoulême
Agathoxylon, Berriasien. Musée d'Angoulême
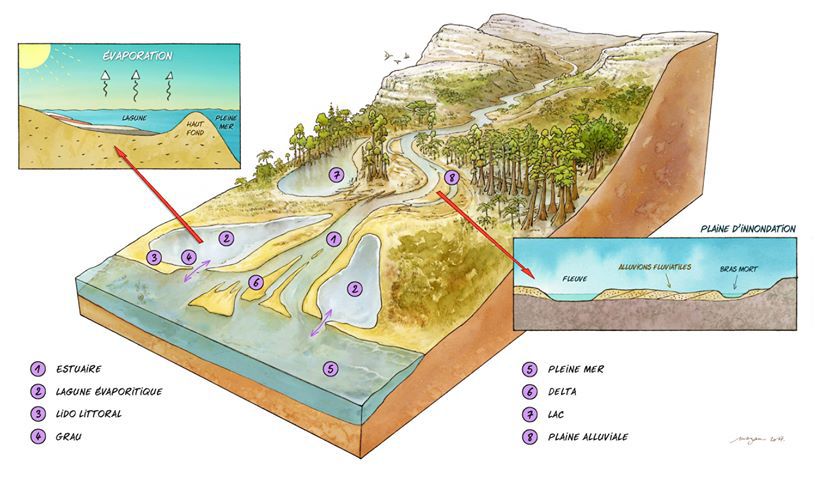
Éléments paléoenvironnementaux.
Les fossiles découverts et l’étude des sédiments permettent de se faire une idée de l’environnement à Cherves.
Ainsi les 3 types de bivalves découverts (Protocardia intexta, Corbula inflexa, et un pectinidae indéterminé) sont connus pour être euryhalines (c’est-à-dire capables de supporter de grandes différences de salinité de l’eau), tandis que les gastéropodes (principalement Viviparus, hydrobiidae, Unio) indiquent un milieu dulçaquicole à saumâtre. La faible diversité de cette faune suggère des conditions variables, de dulçaquicoles à oligohalines.
Les charophytes indiquent un milieu d’eau saumâtre. Les restes retrouvés indiquent soit un environnement de dépôt dulçaquicole, impliquant des conditions saumâtres dans des zones voisines, soit la prédominance d’un milieu saumâtre pendant certaines périodes (Colin et al. 2004, El Albani et al. 2004).
L’étude des palynomorphes montrent une prédominance des variétés terrestres (plus de 90% des restes). Dans le niveau C7 une incursion marine a été mise en évidence par la présence de palynomorphes marins. Cette étude a aussi permis de déterminer des conditions climatiques globalement sèches, surtout dans la partie supérieure de la zone étudiée.

Une étude des fragments d’œufs a permis de déterminer 4 types correspondant à 2 tortues, un crocodylomorphe et un dinosaure (2003, Garcia). En 2008, une nouvelle étude (Grellet-Tinner et al.) a donné 4 types de coquilles : une de lézard (ce qui en ferait la plus ancienne coquille d’œuf de squamate connue), une de crocodylomorphe, une de tortue et une de théropode.
Ces fragments sont de tous de taille millimétrique, avec des angles émoussés (ce qui indique un transport abrasif), et présentant un piquetage en surface (conséquence d’un séjour en milieu acide, probablement en environnement stagnant).
Les coquilles de crocodylomorphe et de lézard présentent des nodules en surface, ce qui indiquerait un lieu de ponte milieu humide.
Les fragments non traités à l’eau oxygénée présentent sur leur face interne un réseau serré de très longs filaments de 20 à 50 nm de diamètre émergeant de la structure cristalline, ces filaments sont interprétés comme étant un réseau de fibres protéiques (Grellet-Tinner et al. 2008). La conservation de telles structures indique un enfouissement rapide ou une stagnation des eaux associée à des bactéries et une probable anoxie du milieu de stagnation, favorable à la conservation. Ces fibres protéiques, qui seraient des fibres de collagènes, représenteraient les plus anciennes protéines connues.
Schnyder (2003) a mis en évidence la faible présence de matière organique très fortement altérée.
Les faciès évaporitiques à la base de la série présentent des teneurs en carbone organique légèrement plus élevées, ce qui indiquerait le confinement du milieu favorable à la préservation de la matière organique ainsi qu’à la prolifération de certains types d’organismes. Leur faible teneur en carbone organique indique une altération rapide. Le milieu devait être suffisamment oxygéné, correspondant peut-être à une tranche d’eau trop faible pour qu’une couche anoxique ou disoxyque se mette en place.
La seconde partie de la couche ne présentait pas de conditions favorables à la conservation de matière organique. Ce qui pourrait correspondre à une sédimentation plus carbonatée, des émersions probablement plus fréquentes et un milieu plus ouvert et oxygéné.
8 lithofaciès, associés à des environnements de dépôts, ont été reconnu par El Albani et al. (2004).
- lithofaciès 1, consistant en un gypse stromatolitique avec laminations cryptomicrobiennes, sans fentes de dessiccation ni érosion. Cette formation est supposée subaquatique, représantative d’un environnement de sebkha .
- lithofaciès 2, réunissant des bancs de gypse décimétriques composés de cristaux orientés aléatoirement, avec présence de nodules de sulfates, laminations cryptomicrobiennes, peloïdes épars, rides et débris de bois. Ces dépôts caractériseraient un environnement peu profond (quelques mètres) de lagune ou plate-forme interne.
- lithofaciès 3, se composant de marne dolomitique gypsifère, avec laminations millimétriques. Ce qui traduirait un environnement peu profond de plate-forme interne ou de lagune, sous influence tidale.
- lithofaciès 4, se composant d’argile dolomitique indurée avec intercalations de couches bioturbées formées d’argile silteuse verte. On retrouve dans ces dépôts des valves d’ostracodes et des écailles de poissons. Ces dépôts ont été mis en place dans un milieu peu profond d’eau saumâtre, subissant une forte évaporation.
- lithofaciès 5, défini par un calcaire beige de plusieurs centimètres, incluant pelotes fécales, galets, mous, ostracodes, bivalves et fragments de vertébrés. Tout cela indique un environnement de transition entre shoreface supérieur et inférieur.
- lithofaciès 6, consistant en un calcaire intercalé avec des remplissages marno-calcaires. Il n’est connu que dans la zone à forte accumulation de macro-restes de vertébrés. Le contenu biologique est complété par des charophytes, ostracodes, gastéropodes et bivalves. Il est interprété comme un environnement de foreshore à shoreface, alimenté par des flux périodiques typiques d’une tempête.
- lithofaciès 7, constitué d’un calcaire oolithique, présentant des surfaces de réactivation. Il renferme des coquilles d’ostracodes, bivalves et gastéropodes. Ce dépôt a été interprété comme le témoin d’une barre séparant l’environnement sous influence marine de l’environnement de sebkha du lithofaciès 1.
- lithofaciès 8, caractérisé par des marnes noires, finement laminées, montrant des signes de bioturbation et renfermant des substances organiques. Il renferme ostracodes, échinodermes, restes de poissons et palynomorphes marins.
Ces constations permettent de proposer l’environnement de dépôt suivant.
L’unité 1, caractérisée par deux types d’environnements : un environnement hypersalin type sebkha et un environnement de plate-forme interne à lagune, en partie confiné. Ces environnements ont pu intégrer occasionnellement des conditions marines.
L’unité 2 débute par des dépôts en eau saumâtre, faisant la transition avec les gypses sous-jacents. Les conditions varient de euryhaline à la base de la série à dulçaquicole au sommet, ce qui traduit une influence continentale croissante.
Les évaporites permettent aussi de nous faire une idée sur l’environnement de Cherves au Berriasien. Le gypse est de pureté différente suivant le niveau où il se trouve, le plus profond est le plus impur (forte présence d’argiles intercalées et contient plus de 5% de chlore), tandis que le banc le plus épais est lui le plus pur.
Platel (1980) indique que pour la formation de sédiments gypseux ou anhydriques, la genèse la plus courante est la précipitation directe des saumures. Elles sont piégées par densité dans des bassins lagunaires dont le fond est isolé de la mer ouverte. Mais pour pouvoir former de grandes quantités d’évaporites il est nécessaire qu’il y ait une alimentation périodique en eau de mer.
Dans un système de lagune les apports d’eau peuvent provenir des précipitations, de la mer, d’apports fluviatiles ou du sous-sol, ce qui influera sur le type d’évaporite. Dans le cas de Cherves-de-Cognac, il est probable que l’eau de mer ait fourni la majeure partie des sulfates nécessaires à la précipitation du gypse, le calcium impliqué pouvant provenir d’une autre source. S’il n’y avait eu que l’eau de mer, les dépôts évaporitiques n’auraient été principalement composé que d’halite (Lagny et al. 2001, Schnyder, com. pers.).
Les alternances entre les lamines argileuses et les bancs de gypse sont interprétées comme des alternances intertidal/supratidal, les lamines correspondant à un apport d’eau supérieur à l’évaporation et inversement pour les dépôts évaporitiques (West, 1975). Plusieurs facteurs peuvent expliqués ces cycles : fluctuations eustatiques, évaporation de la colonne d’eau, progradation corrélée au remplissage du lagon par les sédiments entre deux apports d’eau importants (Schnyder, com. pers.).
Différentes interprétations de l’environnement ont été proposées : Coquand, 1858 ; Delfaud et Gottis, 1966 ; Platel, 1980 ; Bourgueil et al., 1986 ; Schnyder, 2003 ; El Albani, 2004.
D’après les dernières études (Pouech, 2008), on peut donner l’interprétation suivante :
-la partie basale avec des conditions saumâtres, enregistrant une incursion marine (couche C7).
-la partie inférieure de la seconde unité est soumise à des conditions et des salinités variables, dans un milieu faiblement anoxique.
-la partie sommitale enregistre une accentuation progressive des conditions sèches, que l’on peut associer à un milieu plus ouvert, plus oxygéné, à des émersions plus fréquentes, enregistrant des influences dulçaquicoles.
-les différents micro-fossiles indiquent un faible transport.
L’environnement de dépôt peut être envisagé comme un milieu de type sebkha à la base de série, pour évoluer vers une lagune d’eau douce à saumâtre, voire un milieu lacustre dans la partie sommitale.
Les fossiles découverts à Cherves de Cognac proviennent pour la plus grande majorité de 3 niveaux consécutifs (C34, C35, C36), et il s’agit d’espèces à influence continentale et non maritime. Le dépôt a eu lieu sur une grande surface (plusieurs hectares) et rares sont les éléments en connexion anatomique, les restes ont une bonne qualité de conservation mais ils sont souvent abîmés, déformés par la compaction des marnes. Les fossiles n’ont aucune usure liée à un transport (par exemple, par un cours d’eau) et ne sont pas réparties suivant une orientation précise, ce qui indique qu’ils n’ont pas été soumis à un courant lors de leur dépôt. Certains ont des cassures nettes ou une position incompatible avec un dépôt calme (positionnement vertical).
On peut déduire de tout cela que la majorité des restes fossiles a été soumis à un transport rapide mais violent depuis les terres émergées.
Tous ces éléments permettent d’affirmer que le gisement de Cherves de Cognac s’est formé suite à un événement violent et brusque. Glissement de terrain ? Inondation ? Tempête ? Tsunami ?
Biodiversité du début Crétacé
Outre l’étude quantitative et qualitative des restes fossiles, ce gisement a permis d’énormes progrès dans l’étude alimentaire et du paléoenvironnement.
Pour l’approche alimentaire l’importante quantité de restes dentaires a permis une étude quantitative des traces de micro-usure dentaire. Les aliments endommageant l’émail des dents, sous la forme de stries et de petits éclats, il est possible d’analyser ces dégâts afin de découvrir le régime alimentaire de ces animaux disparus. La forme, la taille et la fréquence de ces dégâts permettent de déterminer dureté et type d’aliment, tandis que le nombre, la proportion relative et leur orientation permettent de déterminer le régime alimentaire.
Pour arriver à ces résultats il faut utiliser d’importants moyens, microscopie électronique à balayage, traitement numérique des photos, calcul statistique, reconstitution 3D micrométrique…
Pour l’approche paléoenvironnementale, il a été fait appel à la biogéochimie isotopique de l’oxygène.
Tous les constituants des organismes vivants contiennent les 2 isotopes stables de l’oxygène (18O et 16O), pour les fossiles ces isotopes sont très bien conservés dans les tissus fortement minéralisés, tels les os, mais surtout l’émail dentaire.
Leur proportion relative dépend de la température et est influencée par la salinité, ce qui permet donc de connaitre avec une grande précision les conditions environnementales lors de la formation de ces fossiles.
Ces études ont permis de déterminer que l’environnement était globalement chaud et aride.
Pour terminer, le Synchrotron de Grenoble, un puissant accélérateur de particules, a permis d’obtenir des renseignements sur le taux de croissance et la physiologie des vertébrés de Cherves.
Tous ces éléments font de Cherves-de-Cognac un gisement d’exception et de nombreuses études sont toujours en cours, ce qui fait que de nouvelles découvertes sont encore possibles.
Au nombre de ces études il reste, entre autre :
-les micro-restes, plus de 18 tonnes de sédiments ayant été traités, il reste encore à trier et faire une étude quantifiée pour comprendre précisément les modalités de dépôts et l’organisation de l’assemblage faunistique.
-les mammifères, étendre l’étude de la biodiversité comparée à l’échelle continentale, voire mondiale.
-la mise en place de la thermorégulation chez les mammifères, possible grâce à l’évolution des techniques.
-la description des nouvelles espèces.
Une partie des fossiles découverts est visible au musée d’Angoulême
http://musee-angouleme.fr/
Et merci à Mazan pour les illustrations.
Bibliographie.
-BILLON-BRUYAT J.-P., 2003: Les écosystèmes margino-littoraux du Jurassique terminal et du Crétacé basal d'Europe occidentale: biodiversité, biogéochimie et l'événement biotique de la limite Jurassique-Crétacé. Thèse soutenue le 05 décembre 2003 à l'Université de Poitiers.
-BILLON-BRUYAT J.-P., JM MAZIN & J POUECH, 2010., A stegosaur teeth (Dinosauria, Ornithischi) from the Early Cretaceous of southwestern France.
-BOURGUEUIL B., HANTZPERGUE P. et MOREAU P., 1986 : Carte géologique de la France à 1/50 000, feuille Matha. Bureau de Recherches Géologiques et Minières, Orléans.
-BUFFETAUT E., POUIT D., RIGOLLET L. et ARCHAMBEAU J.-P.,1989: Poissons et reptiles continentaux du Purbeckien de la région de Cognac (Charente). Bull. Soc. géol. Fr. (8), 5, p. 1065-1069.
-COLIN J.-P., EL ALBANI A., FURSICH F. T., MARTIN-CLOSAS C., MAZIN J.-M. et BILLON-BRUYAT J.-P., 2004 : Le gisement “Purbeckien“ de vertébrés de Cherves-de-Cognac, Charente (SW France) : nouvelles données biostratigraphiques. Comptes Rendus Palevol, 3 : 9-16.
- COQUAND H., 1862: Description Physique, Géologique, Paléontologique et Minéralogique du département de la Charente, 420 pages. Tome 2
- COQUAND H., 1858: Description Physique, Géologique, Paléontologique et Minéralogique du département de la Charente, 420 pages. Tome 1
-DELFAUD J. et GOTTIS M., 1966 : Sur quelques figures de sédimentation dans le Portlandien du Lot et sur leur paléogéographique en Aquitaine septentrionale. Actes de la Société Linéenne de Bordeaux, 103(7).
-EL ALBANI A., FURSICH F. T., COLIN J.-P., MEUNIER A., HOCHULI P., MARTIN-CLOSAS M., MAZIN J.-M. et BILLON-BRUYAT J.-P., 2004: Paleoenvironmental reconstruction of the basal Cretaceous vertebrate bearing beds in the Northern part of the Aquitanian Basin (SW France): sedimentological and geochemical evidence. Facies, 50 (2), p.195-215.
-GABILLY J. et al, 1978 : Guides géologiques régionaux : Poitou, Vendée, Charentes.
-GRELLET-TINNER G., POUECH J. et MAZIN J.-M., 2008 : Exquisitely preserved reptile eggshell fragments from the Berriasian site of Cherves-de-Cognac (Charente) : paleobiogical implications. Documents des Laboratoires de Geologie de Lyon, Mid-Mesozoic Life and Environments, 164 : 46-49.
-LAGNY P., ROUCHY J.-M., ORSZAG-SPERBER F., PIERRE C. et GUILHAMOU N., 2001 : Les évaporites, dossier. Géochronique, 80.
-LE LOEUFF J., BUFFETAUT E. and MERSER C., 1996: Découverte d'un dinosaure sauropode tithonien dans la région de Cognac (Charente). Géologie de la France, n°2, 1996, pp. 79-81.
-MAZIN J.-M., BILLON-BRUYAT J.-P., POUECH J. and HANTZPERGUE P., 2006: The Purbeckian site of Cherves de- Cognac (Berriasian, Early Cretaceous, southwest France): a continental ecosystem accumulated in an evaporitic littoral depositional environment. 9th International Symposium on Mesozoic Terrestrial Ecosystems and Biota, Abstracts and Proceedings Volume 84-88.
-MAZIN J.-M., BILLON-BRUYAT J.-P. and POUECH ., 2005: The Purbeckian crocodilians from Cherves-De-Cognac (Berriasian, southwestern France). 53rd Symposium of Vertebrate Palaeontology and Comparative Anatomy, 2005 (London).
-MAZIN J.-M. and BILLON-BRUYAT J.-P., 2003: Nouveau dinosaure à Cognac. Pour La Science, n°306 (avril 2003).
-MAZIN J.-M. and BILLON-BRUYAT J.-P., 2002: Le gisement paléontologique de Cherves-de-Cognac. Pour La Science, n°293 (mars 2002), p.30-31.
-MAZIN J.-M., BILLON-BRUYAT J.-P., LENGLET T and TOURNEPICHE J.-F., 2001: New excavation on the "Purbeckian" site of Cherves (south-western France): a rich reptilian community revealing a terrestrial ecosystem. 6th European Workshop on Vertebrate Palaeontology - Florence and Montevarchi (Italy) - September 19-22, 2001.
-PLATEL J.-P., 1980 : Le bassin gypsifère des Charentes. Service Géologique Régional Poitou-Charentes, Poitiers, France, 47pp.
-POUECH J., MAZIN J.-M., TAFFOREAU P., 2008: The site of Cherves-de-Cognac (Berriasian, SW France): a case of "discrete" Konzervat Lagerstätt leading to physiological conclusions. Geophysical Research Abstracts, Vol. 10, EGU General Assembly 2008.
-POUECH J., MAZIN J.-M. and BILLON-BRUYAT J.-P., 2006: Microvertebrate biodiversity from Cherves-de-Cognac (Lower Cretaceous, Berriasian: Charente, France). 9th International Symposium on Mesozoic Terrestrial Ecosystems and Biota, Abstracts and Proceedings Volume 96-100.
-POUECH J. and MAZIN J.-M, 2006: Taxonomic diversity of micro-vertebrates from Cherves-de-Cognac (Berriasian, Charente, France). 54th Symposium of Vertebrate Palaeontology and Comparative Anatomy, 12-16/09/2006 (Paris).
-POUECH J., 2005: Mammals from Cherves-de-Cognac (Charente, France, Berriasian). 53rd Symposium of Vertebrate Palaeontology and Comparative Anatomy, 2005 (London).
-POUECH J., 2008: Position des mammifères dans les écosystèmes mésozoïques d'Europe.
-POUECH J., 2004: Les micro-restes de vertébrés de Champblanc (Berriasien, Cherves-de-Cognac, Charente). Quantification de la biodiversité. Mémoire DEA, Université de Poitiers - Laboratoire de Paléontologie, 37p.
-SCHNYDER J., 2003: Le passage Jurassique/Crétacé : Evénements instantanés, variations climatiques enregistrées dans les faciès purbeckiens français (Boulonnais, Charentes) et anglais (Dorset). Comparaison avec le domaine téthysien. Thèse soutenue le 05 juillet 2003 à l'Université de Lille 1.
-VULLO R., NERAUDEAU D. and LENGLET T., 2007: Dinosaur teeth from the Cenomanian of Charentes, western France: evidence for a mixed Laurasian-Gondwanan assemblage. Journal of Vertebrate Paleontology, Vol.27, issue 4, p.931-943.
-WEST I. 1975 : Evaporites and associated sediments of the basal Purbeck Formation (Upper Jurassic) of Dorset. Proceedings of the Geologists Association 86(2) : 202-225.
« Dossier sur le gisement paléontologique de Cherves-de-Cognac (carrière de Champblanc) »
(http://dinonews.net/dossiers/gisements/cherves.php5), sur dinonews.net
Les albums-photos à la une !
Le Burdigalien inférieur de Gironde

Album 1
Album
2
Le MHN de La Rochelle

Article +
Album
Le gisement paléontologique d'Angeac

Article + Album
Le MNHN de Paris

Article + Album
Une manifestation, une exposition, une publication, une découverte en rapport avec la paléontologie et les Charentes ? Envoyez nous vos communiqués à :
Devenez acteur de ce site-web !
Vous êtes passionné de paléontologie ? Vous avez écrit un article sur la paléontologie des Charentes et vous ne savez pas où le publier ? Devenez acteur de ce site-web en nous proposant vos articles !